
Tehnologia ADN recombinat
Tehnologia ADN recombinat
Tehnologia ADN-ului recombinat este cunoscută şi sub numele de clonarea genelor sau clonare moleculară. Ea reprezintă termenul generic sub care sunt reunite un numar de proceduri experimentale care conduc către transferul de informație genetică reprezentată de acidul dezoxiribonucleic – AND, de la un organism la altul.
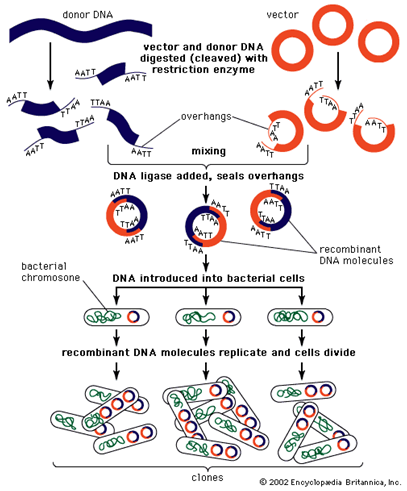
Fenomenul ADN-ului recombinat poate fi sintetizat în umatoarele etape:
• ADN-ul (ADN-ul clonat, ADN-ul inserat, ADN-ul țintă, ADN-ul străin) extras de la un organism donator, este scindat din punct de vedere enzimatic (taiat, incorporat), reunitși legat cu o altă entitate ADN (un vector de clonare) în vederea formării unei molecule noi de ADN recombinat (vector inserat de clonare ADN construit, ADN construit);
• acest ADN construit este transferat în interior și păstrat în cadrul unei celule gazdă. Introducerea ADN-ului în interiorul unei celule bacteriene gazdă, poartă denumirea de transformare;
• aceste celule gazdă care preiau ADN-ul construit (celulele transformate) sunt identificate şi selectate (separate, izolate) de acelea care nu iau parte la acest proces (nu procedează în același mod);
• dacă este necesar, un ADN construit poate fi preparat în aşa fel încât producerea de proteină care se gasește în codul secvenței de ADN clonat, este realizată de către celula gazdă.
Tehnologia ADN-ului recombinat s-a dezvoltat ca urmare a cercetărilor făcute în cadrul biologiei moleculare, studiului enzimologiei acidului nucleic,și a geneticii moleculare atât a virușilor bacterieni, câtși a elementelor bacteriene extracromozomiale ale ADN (plastidele). Cu toate acestea, tehnologia ADN-ului recombinat n-ar fi putut exista fără validitatea enzimelor (enzimele de restricție, endonucleaze de restrictie) care identifică secvențele de ADN specific dublu catenar și care desparte ADN-ul, atât în catene câtși în acele secvențe.
În ceea ce privește clonarea moleculară, atât sursa ADN care conţine secvenţa ţintă, cât şi vectorul de clonare trebuie sa fie secţionate fără excepţie în fragmente fine reproductibile. Izolarea cromozomială ADN este realizată fie prin pătrunderea unui ac de calibru foarte mic sau prin procedeul sonic. Oricare din cele doua metode trebuie să aibă ca urmare producerea de fragmente de ADN sub forma deșiruri, care pot varia în lungime de la 0,3 la 5 kilobase (kb). Din nefericire, aceste proceduri uzuale pot induce ruperi pe alocuri, așa încât ori de cate ori o mostră de ADN este tratată, pot fi generateși alte seturi de fragmente. Numai în cazul enzimelor bacteriene care secționează în interior moleculele de ADN în secvențe catenare perechi, s-a evidențiat că întradevăr clonarea moleculară este viabilă. Aceste enzime poartă numele de endonucleaze de restricție de tip II.
Unul din primele tipuri de endonucleaze de restricție evidențiate au fost cele de la bacteria Escherichia Coli si a primit denumirea de EcoRI. Această enzimă se leaga de o zonă ADN cu o secvență specifică palindromică (cele douășiruri sunt identice în această zonă atunci când indică aceeași polaritate, ex. de la 5' la 3') din cele 6 perechi (bp), secționandu-se între reziduurile de guaninăși adenină ale ambelorșiruri. Bifurcația are loc între legătura nucleotidică dintre oxigenși carbon 3' a particulei de zahar a unei nucleotideși grupul de fosfați, care se află atașat de carbon 5' din particula de zahăr a nucleotidei adiacente. Această bifurcație de o simetrie surprinzătoare a ADN-ului EcoRI produce douășiruri singulare, complementare la capete, fiecare având câte patru nucleotide cași extensii. În acest caz, fiecare extensie așirului se termină într-un grup 5' fosfat, iar grupul 3'-hidroxil de lașirul opus este absent.
În completare la EcoRI, sute de alte endonucleaze de restricție de tip II au fost izolate de la diverse bacterii. Protocolul de denumire pentru aceste enzime este același cu cel pentru EcoRi; genul este evidențiat cu litere mari, iar primele două litere ale numelui speciei sunt reprezentate cu litere mici. Desemnareașirului este adesea scăpată din vedere în ceea ce privește denumirea, fiind utilizate cifrele romane pentru a desemna ordinea caracterizării diferitelor endonucleaze de restricție ale aceluiași organism. De exemplu, HpaI si HpaII reprezintă primulși al doilea tip de endonucleaze de restricție care au fost izolate de la Haemophilus parainfluenzae.
Cele mai multe dintre secvenţele palindromice fac parte din tipul II de endonucleaze de restricție tăiate şi legate de molecula de ADN, așa numitele situsuri de recunoaștere. Unele dintre endonucleazele de restricție ale ADN-ului încorporat (scindat), lasă extensiile 5' fosfat, altele lasă extensiile 3' hidroxil şi altele taie în structura scheletului ambele catene din cadrul situsului de recunoaștere pentru a produce molecule de ADN. Lungimea situsurilor de recunoaştere a diferitelor enzime poate fi patru, cinci, şase, opt sau mai multe perechi de nucleotide. Datorită frecvenței cu care situsurile de recunoaștere apar în ADN, endonucleazele de restricție care scindează în cadrul situsului de patru (divizat în patru), de şase (divizat în şase) perechi de nucleotide sunt utilizate în cele mai multe ori în clonarea moleculară.
Sursa:
1. Antohi, Şt., Gavrilă, L., 1981 - Progrese în genetica moleculară. Editura Ştiiţifică şi Enciclopedică, Bucureşti
2. Gavrilă, L., Dabala, I., 1975 - Descifrând tainele eredităţii. Editura Dacia, Cluj Napoca
3. Watson, J.D., Crick, F.H.C., 1953 - The structure of DNA, Cold Spring Harbor Symposium, Quant. Biol., 18
4. Wilkins. M.H.F., 1963 - The molecular strucutre of DNA, J. Chim. Phzs. 58
August, 2011
Ing. Lucian Ciprian MELUŢ